Значение симпатической и парасимпатической нервной системы. Вегетативная нервная система: парасимпатическая иннервация и ее нарушения
Парасимпатическая нервная система осуществляет сужение бронхов, замедление и ослабление сердечных сокращений; сужение сосудов сердца; пополнение энергоресурсов (синтез гликогена в печени и усиление процессов пищеварения); усиление процессов мочеобразования в почках и обеспечение акта мочеиспускания (сокращение мышц мочевого пузыря и расслабление его сфинктера) и др. Парасимпатическая нервная система преимущественно оказывает пусковые влияния: сужение зрачка, бронхов, включение деятельности пищеварительных желез и т. п.
Деятельность парасимпатического отдела вегетативной нервной системы направлена на текущую регуляцию функционального состояния, на поддержание постоянства внутренней среды - гомеостаза. Парасимпатический отдел обеспечивает восстановление различных физиологических показателей, резко измененных после напряженной мышечной работы, пополнение израсходованных энергоресурсов. Медиатор парасимпатической системы - ацетилхолин, снижая чувствительность адренорецепторов к действию адреналина и норадреналина, оказывает определенное антистрессорное влияние.
Рис. 6. Вегетативные рефлексы
Влияние положения тела на частоту сердечных сокращений
(уд./мин). (По.МогендовичМ.Р., 1972)
3.6.4. Вегетативные рефлексы
Через вегетативные симпатические и парасимпатические пути ЦНС осуществляет некоторые вегетативные рефлексы, начинающиеся с различных рецепторов внешней и внутренней среды: висцеро-висцеральные (с внутренних органов на внутренние органы - например, дыхательно-сердечный рефлекс); дермо-висцеральные (с кожных покровов - изменение деятельности внутренних органов при раздражении активных точек кожи, например, иглоукалыванием, точечным массажем); с рецепторов глазного яблока - глазо-сердечный рефлекс Ашнера (урежение сердцебиений при надавливании на глазные яблоки - парасимпатический эффект); моторно-висцеральные- например, ортостэтическая проба (учащение сердцебиения при переходе из положения лежа в положение стоя - симпатический эффект) и др. (рис. 6). Они используются для оценки функционального состояния организма и особенно состояния вегетативной нервной системы (оценки влияния симпатического или парасимпатического ее отдела).
11. ПОНЯТИЕ О НЕРВНО МЫШЕЧНОМ(ДВИГАТЕЛЬНОМ) АППАРАТЕ. ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ(ДЕ) И ИХ КЛАССИФИКАЦИЯ. ФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ РАЗЛИЧНЫХ ТИПОВ ДЕ И ИХ КЛАССИФИКАЦИЯ. ФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ РАЗЛИЧНЫХ ТИПОВ ДЕ.(ПОРОГ АКТИВАЦИИ,СКОРОСТЬ И СИЛА СОКРАЩЕНИЯ, УТОМЛЯЕМОСТЬ И ДР) Значение типа ДЕ при различных видах мышечной деятельности.
12. Мышечная композиция. Функциональные возможности разных типов мышечных волокон (медленные и быстрые). Их роль в проявлении мышечной силы, скорости и выносливости. Одной из важнейших характеристик скелетных мышц, влияющих на силу сокращения, является состав (композиция) мышечных волокон. Различают 3 типа мышечных волокон - медленные неутомляемые (I типа), быстрые неутомляемые или промежуточные (11-а типа) и быстрые утомляемые (11-б типа).
Медленные волокна (1 типа), их обозначают также SO - Slow Oxydative (англ. - медленные окислительные) - это выносливые (неутомляемые) и легко возбудимые волокна, с богатым кровоснабжением, большим количеством митохондрий, запасов миоглобина и
с использованием окислительных процессов энергообразования (аэробные). Их, в среднем, у человека 50%. Они легко включаются в работу при малейших напряжениях мышц, очень выносливы, но не обладают достаточной силой. Чаще всего они используются при поддержании ненагрузочной статической работы, например, при сохранении позы.
Быстрые утомляемые волокна (11-б типа) или FG - Fast Glicolitic (быстрые гликолитические) используют анаэробные процессы энергообразования (гликолиз). Они менее возбудимы, включаются при больших нагрузках и обеспечивают быстрые и мощные сокращения мышц. Зато эти волокна быстро утомляются. Их примерно 30%. Волокна промежуточного типа (П-а) - быстрые неутомляемые, окислительные, их около 20%. В среднем, для разных мышц характерно различное соотношение медленных неутомляемых и быстрых утомляемых волокон. Так, в трехглавой мышце плеча преобладают быстрые волокна (67%) над медленными (33%), что обеспечивает скоростно-силовые возможности этой мышцы (рис. 14), а для более медленной и выносливой камбаловидной мышцы характерно наличие 84% медленных и всего 16% быстрых волокон (Салтан Б., 1979).
Однако, состав мышечных волокон в одной и той же мышце имеет огромные индивидуальные различия, зависящие от врожденных типологических особенностей человека. К моменту рождения человека его мышцы содержат лишь медленные волокна, но под влиянием нервной регуляции устанавливается в ходе онтогенеза генетически заданное индивидуальное соотношение мышечных волокон разного типа. По мере перехода от зрелого возраста к пожилому число быстрых волокон у человека заметно снижается и, соответственно, уменьшается мышечная сила. Например, наибольшее количество быстрых волокон в наружной головке 4-х главой мышцы бедра мужчины (около 59-63%) отмечается в возрасте 20-40 лет, а в возрасте 60-65 лет их число почти на 1/3 меньше (45%).

Рис. 14. Состав мышечных волокон в разных мышцах
Медленные - черным цветом; быстрые - серым
Количество тех или других мышечных волокон не изменяется в процессе тренировки. Возможно только нарастание толщины (гипертрофия) отдельных волокон, а также некоторое изменение свойств промежуточных волокон. При направленности тренировочного процесса на развитие силы происходит нарастание объема быстрых волокон, что и обеспечивает повышение силы тренируемых мышц.
Характер нервных импульсов изменяет силу сокращения мышц тремя способами:
Существенное значение имеют механические условия работы мышцы -точка приложения ее силы и точка приложения сопротивления (поднимаемого груза). Например, при сгибании в локте вес поднимаемого груза может быть порядка 40 кг и более, при этом сила мышц-сгибателей достигает 250 кг, а тяга сухожилий - 500 кг.
Между силой и скоростью сокращения мышцы существует определенное соотношение, имеющее вид гиперболы (соотношение сила - скорость, по А. Хиллу). Чем выше сила, развиваемая мышцей, тем меньше скорость ее сокращения, и наоборот, с нарастанием скорости сокращения падает величина усилия. Наибольшую скорость развивает мышца, работающая без нагрузки. Скорость мышечного сокращения зависит от скорости передвижения поперечных мостиков, т. е. от частоты гребковых движений в единицу времени. В быстрых ДЕ эта частота выше, чем в медленных ДЕ, и, соответственно, потребляется больше энергии АТФ. Во время сокращения мышечных волокон в 1 с происходит примерно от 5 до 50 циклов прикрепления-отсоединения поперечных мостиков. При этом никаких колебаний силы в целой мышце не ощущается, так как ДЕ работают асинхронно. Лишь при утомлении возникает синхронная работа ДЕ, и в мышцах появляется дрожь (тремор утомления).
13. ОДИНОЧНОЕ И ТЕТАНИЧЕСКОЕ СОКРАЩЕНИЕ МЫШЕЧНОГО ВОЛОКНА. ЭЛЕКТРОМИОГРАММА. При единичном надпороговом раздражении двигательного нерва или самой мышцы возбуждение мышечного волокна сопровождается
одиночным сокращением. Эта форма механической реакции состоит из 3 фаз: латентного или скрытого периода, фазы сокращения и фазы расслабления. Самой короткой фазой является скрытый период, когда в мышце происходит электромеханическая передача. Фаза расслабления обычно в 1.5-2 раза более продолжительна, чем фаза сокращения, а при утомлении затягивается на значительное время.
Если интервалы между нервными импульсами короче, чем длительность одиночного сокращения, то возникает явление суперпозиции - наложение механических эффектов мышечного волокна друг на друга и наблюдается сложная форма сокращения - тетанус. Различают 2 формы тетануса - зубчатый тетанус, возникающий при более редком раздражений, когда происходит попадание каждого следующего нервного импульса в фазу расслабления отдельных оди ночных сокращений, и сплошной или гладкий тетанус, возникающий при более частом раздражении, когда каждый следующий импульс попадает в фазу сокращения (рис. 11). Таким образом, (в некоторых границах) между частотой импульсов возбуждения и амплитудой сокращения волокон ДЕ существует определенное соотношение: при небольшой частоте (например, 5-8 имп. в 1с)

Рис. П. Одиночное сокращение, зубчатый и сплошной тетанус камбаловидпой мышцы человека (по: Зимкин Н.В. и др., 1984). Верхняя кривая - сокращение мышцы, нижняя - отметка раздражения мышцы, справа указана частота раздражени я
возникают одиночные сокращения, при увеличении частоты (15-20 имп. в 1с) - зубчатый тетанус, при дальнейшем нарастании частоты (25-60 имп. в 1 с) - гладкий тетанус. Одиночное сокращение - более слабое и менее утомительное, чем тетаническое. Зато тетанус обеспечивает в несколько раз более мощное, хотя и кратковременное сокращение мышечного волокна.
Сокращение целой мышцы зависит от формы сокращения отдельных ДЕ и их координации во времени. При обеспечении длительной, но не очень интенсивной работы, отдельные ДЕ сокращаются попеременно (рис. 12), поддерживая общее напряжение мышцы на заданном уровне (например, при беге на длинные и сверхдлинные дистанции). При этом отдельные ДЕ могут развивать как одиночные, так и тетанические сокращения, что зависит от частоты нервных импульсов. Утомление в этом случае развивается медленно, так как, работая по очереди, ДЕ в промежутках между активацией успевают восстанавливаться. Однако для мощного кратковременного усилия (например, поднятия штанги) требуется синхронизация активности отдельных ДЕ, т. е. одновременное возбуждение практически всех ДЕ. Это, в свою очередь, требует одновременной активации

Рис. 12. Различные режимы работы двигательных единиц (ДЕ)
соответствующих нервных центров и достигается в результате длительной тренировки. При этом осуществляется мощное и весьма утомительное тетаническое сокращение.
Амплитуда сокращения одиночного волокна не зависит от силы надпорогового раздражения (закон «Все или ничего»). В отличие от этого, при нарастании силы надпорогового раздражения сокращение целой мышцы постепенно растет до максимальной амплитуды.
Работа мышцы с небольшой нагрузкой сопровождается редкой частотой нервных импульсов и вовлечением небольшого числа ДЕ. В этих условиях, накладывая отводящие электроды на кожу над мышцей и используя усилительную аппаратуру, можно на экране осциллографа или с применением чернильной записи на бумаге зарегистрировать одиночные потенциалы действия отдельных Д Е. В случае же значительных напряжений потенциалы действия многих ДЕ алгебраически суммируются и возникает сложная интегрированная кривая записи электрической активности целой мышцы - электромиограмма (ЭМГ).
Форма ЭМГ отражает характер работы мышцы: при статических усилиях она имеет непрерывный вид, а при динамической работе - вид отдельных пачек импульсов, приуроченных, в основном, к начальному моменту сокращения мышцы и разделенных периодами «электрического молчания». Особенно хорошо ритмичность появления подобных пачек наблюдается у спортсменов при циклической работе (рис. 13). У маленьких детей и неадаптированных к такой работе лиц четких периодов отдыха не наблюдается, что отражает недостаточное расслабление мышечных волокон работающей мышцы.
Чем больше внешняя нагрузка и cилa сокращения мышцы, тем выше амплитуда ее ЭМГ. Это связано с увеличением частоты нервных импульсов, вовлечением большего числа ДЕ в мышце и синхронизацией

Рис. 13. Электромиограмма мышц-антагонистов при циклической работе
их активности. Современная многоканальная аппаратура позволяет производить одновременную регистрацию ЭМГ многих мышц на разных каналах. При выполнении спортсменом сложных движений можно видеть на полученных ЭМГ кривых не только характер активности отдельных мышц, но и оценить моменты и порядок их включения или выключения в различные фазы двигательных актов. Записи ЭМГ, полученные в естественных условиях двигательной деятельности, можно передавать к регистрирующей аппаратуре по телефону или радиотелеметрически. Анализ частоты, амплитуды и формы ЭМГ (например, с помощью специальных компьютерных программ) позволяет получить важную информацию об особенностях техники выполняемого спортивного упражнения и степени ее освоения обследуемым спортсменом.
По мере развития утомления при той же величине мышечного усилия амплитуда ЭМГ нарастает. Это связано с тем, что снижение сократительной способности утомленных ДЕ компенсируется нервными центрами вовлечением в работу дополнительных ДЕ, т. е. путем увеличения количества активных мышечных волокон. Кроме того, усиливается синхронизация активности ДЕ, что также повышает амплитуду суммарной ЭМГ.
14. Механизм сокращения и расслабления мышечного волокна. Теория скольжения. Роль саркоплазматического ретикулума и ионов кальция в сокращении. При произвольной внутренней команде сокращение мышцы человека начинается примерно через 0.05 с (50 мс). За это время моторная команда передается от коры больших полушарий к мотонейронам спинного мозга и по двигательным волокнам к мышце. Подойдя к мышце, процесс возбуждения должен с помощью медиатора преодолеть нервно-мышечный синапс, что занимает примерно 0.5 мс. Медиатором здесь является ацетилхолин, который содержится в синаптических пузырьках в пресинаптической части синапса. Нервный им пульс вызывает перемещение синаптических пузырьков к пресинаптической мембране, их опорожнение и выход медиатора в синаптическую щель. Действие ацетилхолина на постсинаптическую мембрану чрезвычайно кратковременно, после чего он разрушается ацетилхолинэстеразой на уксусную кислоту и холин. По мере расходования запасы ацетилхолина постоянно пополняются путем его синтезирования в пресинаптической мембране. Однако, при очень частой и длительной импульсации мотонейрона расход ацетилхолина превышает его пополнение, а также снижается чувствительность постсинаптической мембраны к его действию, в результате чего нарушается проведение возбуждения через нервно-мышечный синапс. Эти процессы лежат в основе периферических механизмов утомления при длительной и тяжелой мышечной работе.
Выделившийся всинаптическую щель медиатор прикрепляется к рецепторам постсинаптической мембраны и вызывает в ней явления деполяризации. Небольшое подпороговое раздражение вызывает лишь местное возбуждение небольшой амплитуды - потенциал концевой пластинки (ПКП).
При достаточной частоте нервных импульсов ПКП достигает порогового значения и на мышечной мембране развивается мышечный потенциал действия. Он (со скоростью 5 ) распространяется вдоль по поверхности мышечного волокна и заходите поперечные
трубочки внутрь волокна. Повышая проницаемость клеточных мембран, потенциал действия вызывает выход из цистерн и трубочек саркоплазматического ретикулума ионов Са, которые проникают в миофибриллы, к центрам связывания этих ионов на молекулах актина.
Под влиянием Садлинные молекулы тропомиозина проворачиваются вдоль оси и скрываются в желобки между сферическими молекулами актина, открывая участки прикрепления головок миозина к актину. Тем самым между актином и миозином образуются так называемые поперечные мостики. При этом головки миозина совершают гребковые движения, обеспечивая скольжение нитей актина вдоль нитей миозина с обоих концов саркомера к его центру, т. е. механическую реакцию мышечного волокна (рис. 10).
Энергия гребкового движения одного мостика производит перемещение на 1 % длины актиновой нити. Для дальнейшего скольжения сократительных белков друг относительно друга мостики между актином и миозином должны распадаться и вновь образовываться на следующем центре связывания Са. Такой процесс происходит в результате активации в этот момент молекул миозина. Миозин приобретает свойства фермента АТФ-азы, который вызывает распад АТФ. Выделившаяся при распаде АТФ энергия приводит к разрушению

Рис. 10. Схема электромеханической связи в мышечном волокне
На А: состояние покоя, на Б - возбуждение и сокращение
да - потенциал действия, мм - мембрана мышечного волокна,
п _ поперечные трубочки, т - продольные трубочки и цистерны с ионами
Са , а - тонкие нити актина, м - толстые нити миозина
с утолщениями (головками) на концах. Зет-мембранами ограничены
саркомеры миофибрилл. Толстые стрелки - распространение потенциала
действия при возбуждении волокна и перемещение ионов Саиз цистерн
и продольных трубочек в миофибриллы, где они содействуют образованию
мостиков между нитями актином и миозином и скольжение этих нитей
(сокращение волокна) за счет гребковых движений головок миозина.
имеющихся мостиков и образованию в присутствии Сановых мостиков на следующем участке актиновой нити. В результате повторения подобных процессов многократного образования и распада мостиков сокращается длина отдельных саркомеров и всего мышечного волокна в целом. Максимальная концентрация кальция в миофибрилле достигается уже через 3 мс после появления потенциала действия в поперечных трубочках, а максимальное напряжение мышечного волокна - через 20 мс.
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает видимая под микроскопом поперечная исчерченность мышцы.
Расслабление мышечного волокна связано с работой особого механизма - «кальциевого насоса», который обеспечивает откачку ионов Саиз миофибрилл обратно в трубочки саркоплазматического ретикулума. На это также тратится энергия АТФ.
15. Механизм регуляции силы сокращения мышц (число активных ДЕ, частота импульсации мотонейронов, синхронизация сокращения мышечных волокон разных ДЕ во времени). Характер нервных импульсов изменяет силу сокращения мышц тремя способами:
1) увеличением числа активных ДЕ - это механизм вовлечения или рекрутирования ДЕ (сначала происходит вовлечение медленных и более возбудимых ДЕ, затем - высокопороговых быстрых Д Е);
2) увеличением частоты нервных импульсов, в результате чего происходит переход от слабых одиночных сокращений к сильным тетаническим сокращениям мышечных волокон;
3) увеличением синхронизации ДЕ, при этом происходит увеличение силы сокращения целой мышцы за счет одновременной тяги всех активных мышечных волокон.
ВНС делится на два отдела - симпатический и парасимпатический. По строению они различаются расположением своих центральных и эффекторных нейронов, своими рефлекторными дугами. Они различаются так же и по своему влиянию на функции иннервируемых структур.
В чем состоят различия этих отделов? Центральные нейроны симпатической нервной системы расположены, как правило, в сером веществе боковых рогах спинного мозга от 8 шейного до 2-3 поясничных сегментов. Таким образом, симпатические нервы всегда отходят только от спинного мозга в составе спинномозговых нервов по передним (вентральным) корешкам.
Центральные нейроны парасимпатической же нервной системы находятся в крестцовых сегментах спинного мозга (2-4 сегменты), но большая часть центральных нейронов находятся в стволе мозга. Большая же часть нервов парасимпатической системы отходят от головного мозга в составе смешанных черепно - мозговых нервов. А именно: из среднего мозга в составе III пары (глазодвигательный нерв) -- иннервируя мышцы ресничного тела и кольцевые мышцы зрачка глаза, из Варолиевого моста выходит лицевой нерв -- VII пара (секреторный нерв) иннервирует железы слизистой оболочки носа, слёзные железы, подчелюстную и подъязычную железы. Из продолговатого мозга отходит IX пара -- секреторный, языкоглоточный нерв, иннервирует околоушные слюнные железы и железы слизистой щек и губ, X пара (блуждающий нерв) -- самая значительная часть парасимпатического отдела ВНС, проходя в грудную и брюшную полости, иннервирует весь комплекс внутренних органов. Нервы, отходящие от крестцовых сегментов (2-4 сегменты), иннервируют органы малого таза и входят в состав подчревного сплетения.
Эффекторные нейроны симпатической нервной системы вынесены на периферию и находятся или в паравертебральных ганглиях (в симпатической нервной цепочке), или превертебрально. Постганглионарные волокна образуют различные сплетения. Среди них наиболее важное значение имеет чревное (солнечное) сплетение, но в его состав входят не только симпатические, но и парасимпатические волокна. Оно обеспечивает иннервацию всех органов расположенных в брюшной полости. Вот почему так опасны удары и травмы верхней части брюшной полости (примерно под диафрагму). Они способны вызвать шоковое состояние.
Эффекторные нейроны парасимпатической нервной системы всегда находятся в стенках внутренних органов (интрамурально). Таким образом, у парасимпатических нервов большая часть волокон покрыты миелиновой оболочкой, и импульсы достигают эффекторных органов быстрее, чем у симпатической. Это обеспечивает парасимпатические нервные влияния, обеспечивающие сбережение ресурсов органа и организма в целом. Внутренние органы, расположенные в грудной и брюшной полости иннервируются главным образом блуждающим нервом (n. vagus), поэтому эти влияния часто называют вагусными (вагальными).
Имеются существенные различия и в их функциональных характеристиках.
Симпатический отдел, как правило, мобилизует ресурсы организма для осуществления энергичной деятельности (усиливается работа сердца, сужается просвет кровеносных сосудов и повышается артериальное давление, учащается дыхание, расширяются зрачки и т.п.), но происходит торможение работы пищеварительной системы, за исключением работы слюнных желез. У животных это происходит всегда (слюна нужна им для зализывание возможных ран), но и у некоторых людей при возбуждении слюноотделение усиливается.
Парасимпатическая, напротив, стимулирует работу пищеварительной системы. Неслучайно после сытного обеда отмечается вялость, нам так хочется поспать. При возбуждении парасимпатической нервная система обеспечивает восстановление равновесия внутренней среды организма. Она обеспечивает работу внутренних органов в состоянии покоя.
В функциональном смысле симпатическая и парасимпатическая системы являются антагонистами, дополняя друг друга в процессе поддержания гомеостазиса, поэтому многие органы получают двойную иннервацию -- и со стороны симпатического, и со стороны парасимпатического отделов. Но, как правило, у разных людей преобладает или тот или другой отдел ВНС. Неслучайно известный отечественный физиолог Л.А. Орбели попытался классифицировать людей по этому признаку. Он выделил три типа людей: симпатикотоники (с преобладанием тонуса симпатической нервной системы) - их отличает сухость кожи, повышенная возбудимость; второй тип -- ваготоники с преобладанием парасимпатических влияний -- для них характерна жирная кожа, замедленные реакции. Третий тип -промежуточный. Из повседневной практики каждый из нас может заметить, что чай и кофе вызывают различную реакцию у людей с разным типом функциональной активности ВНС. Из экспериментов на животных известно, что у животных с разным типом ВНС введение брома и кофеина так же оказывает различные реакции. Но на протяжении жизни человека его тип ВНС может изменяться в зависимости от возраста, периода полового созревания, беременности и других влияний. Несмотря на перечисленные различия, обе эти системы, однако, составляют единое функциональное целое, т. к. интеграция их функций осуществляется на уровне ЦНС. В сером веществе спинного мозга центры вегетативных и соматических рефлексов успешно соседствуют, также как они располагаются близко друг с другом в стволе мозга, и в высших подкорковых центрах. Так же как, в конечном счете, в единстве функционирует вся нервная система.
Функциональное созревание периферических отделов вегетативной нервной системы тесно связано с состоянием высших отделов ЦНС, после рождения, на ранних стадиях постнатального онтогенеза, в основном регулирование осуществляют центры симпатической нервной системы. Тонус парасимпатической системы, в частности блуждающего нерва, отсутствует. Блуждающий нерв включается в рефлекторные реакции на 2-3-м месяце жизни ребенка. Вместе с тем отделы вегетативной нервной системы начинают функционировать в разные сроки онтогенеза неодинаково по отношению различных органов и систем. Так, относительно органов пищеварения сначала включается парасимпатическая система, а симпатическая регуляция начинает действовать в период отнятия младенца от груди. Относительно регуляции деятельности сердца симпатическая система включается раньше вагусной. Как свидетельствуют результаты экспериментальных исследований, передачи возбуждения в вегетативных ганглиях у новорожденных осуществляется адренергическим путем, а не с помощью ацетилхолина, как это наблюдается у взрослых.
Таким образом, симпатичная передача возбуждения в период раннего онтогенеза характеризуется большим количеством адренергических синапсов. В старости симпатические и парасимпатические тонические влияния на деятельность ряда органов ослабевают. Это влияет на ход важных вегетативных реакций и метаболических процессов и тем самым ограничивает адаптивные возможности стареющего организма. Наряду с этим в процессе старения снижается содержание в крови катехоламинов, но повышается чувствительность клеток и тканей к их действия, а также к ряду других физиологически активных веществ. Ослабление вегетативных реакций является одной из причин снижения трудоспособности при старении.
В период старения наступают структурные и функциональные нарушения в вегетативных ганглиях, которые могут препятствовать передаче импульсов в них и влиять на трофику ткани, иннервируется. Существенно изменяется гипоталамическая регуляция вегетативных функций, что является важным механизмом старения организма.
Проекции вегетативных центров представлены и в коре больших полушарий - в основном в лимбической и ростральной части коры. Парасимпатические и симпатические проекции одних и тех же органов проецируются в одни и те же или близко расположенные участки коры, это понятно, т. к. они совместно обеспечивают функции этих органов. Установлено, что парасимпатические проекции в коре представлены гораздо шире, чем симпатические, однако, функционально симпатические влияния более продолжительны, чем парасимпатические. Это связано с различиями медиаторов, которые выделяются окончаниями симпатических (адреналин и норадреналин) и парасимпатических (ацетилхолин) волокон. Ацетилхолин -- медиатор парасимпатической системы - быстро инактивируется ферментом ацетилхолинэстеразой (холинэстеразой) и её влияния быстро сходят на нет, в то время как адреналин и норадреналин инактивируются значительно медленнее (ферментом моноаминоксидазой), их влияние усиливается норадреналином и адреналином, выделяемыми надпочечниками. Таким образом, симпатические влияния длятся дольше и оказываются более выраженными, чем парасимпатические. Однако, во время сна парасимпатические влияния на все наши функции превалируют, что способствует восстановлению ресурсов организма.
Вегетативная нервная система осуществляет два рода рефлексов: функциональные и трофические.
Функциональное влияние на органы состоит в том, что раздражение вегетативных нервов либо вызывает функцию органа, либо тормозит ее («пусковая» функция).
Трофическое влияние состоит в том, что обмен веществ в органах регулируется непосредственно и тем самым определяется уровень их деятельности («корригирующая» функция).
Вегетативные рефлексы по характеру взаимосвязей афферентного и эфферентного звеньев, а также внутрицентральных взаимоотношений принято подразделять на:
- 1) висцеро-висцеральные, когда и афферентное и эфферентное звенья, т.е. начало и эффект рефлекса относятся к внутренним органам или внутренней среде (гастро-дуоденальный, гастрокардиальный, ангиокардиальные и т.п.);
- 2) висцеро-соматические, когда начинающийся раздражением интероцепторов рефлекс за счет ассоциативных связей нервных центров реализуется в виде соматического эффекта. Например, при раздражении хеморецепторов каротидного синуса избытком углекислоты усиливается деятельность дыхательных межреберных мышц и дыхание учащается;
- 3) висцеро-сенсорные, -- изменение сенсорной информации от экстероцепторов при раздражении интероцепторов. Например, при кислородном голодании миокарда имеют место так называемые отраженные боли в участках кожи (зоны Хеда), получающих сенсорные проводники из тех же сегментов спинного мозга;
- 4) сомато-висцеральные, когда при раздражении афферентных входов соматического рефлекса реализуется вегетативный рефлекс. Например, при термическом раздражении кожи расширяются кожные сосуды и суживаются сосуды органов брюшной полости. К сомато-вегетативным рефлексам относится и рефлекс Ашнера-Даньини -- урежение пульса при надавливании на глазные яблоки.
Рефлексы вегетативной нервной системы (симпатической и парасимпатической) условно можно разделить на кожно-сосудистые рефлексы, висцеральные рефлексы, зрачковые рефлексы.
Строение парасимпатического отдела нервной системы. К парасимпатической нервной системе относится комплекс интрамуральных ганглионарных структур, локализованных в стенках внутренних органов, обладающих двигательной активностью (сердце, бронхи, кишечник, матка, мочевой пузырь). Центральные структуры этого отдела расположены в среднем, продолговатом мозге и в крестцовом отделе спинного мозга, а также образованы парасимпатическими нейронами ганглиев, расположенных чаще всего в иннервируемых органах.
В среднем мозге вблизи передних бугров четверохолмия находятся ядра глазодвигательного нерва (III пара черепных нервов). В продолговатом мозге расположены три пары ядер, от которых отходят три пары черепных нервов: лицевой (VII пара), языкоглоточный (IX пара) и блуждающий (X пара). В спинном мозге в боковых рогах трех сегментов крестцовой части локализованы ядра преганг- лионарных парасимпатических нейронов.
Аксоны нейронов среднего мозга направляются к исполнительным органам в составе глазодвигательного нерва; продолговатого мозга - в составе лицевого, языкоглоточного и блуждающего нервов; крестцового отдела спинного мозга - в составе тазовых нервов. Они называются преганглионарными парасимпатическими волокнами.
Из среднего мозга преганглионарные нервные волокна выходят в составе глазодвигательного нерва, проникают через глазную щель в глазницу и заканчиваются на телах постганглионарных нейронов, расположенных в глубине глазницы.
Из продолговатого мозга, из верхнего слюноотделительного ядра преганглионарные волокна идут в составе лицевого нерва (VII пара) и, покидая его, образуют барабанную струну, которая присоединяется к язычному нерву и заканчивается в челюстном или подъязычном ганглии. Его постганглионарные волокна иннервируют подчелюстную слюнную железу.
Из нижнего слюноотделительного ядра продолговатого мозга выходят преганглионарные волокна, вступают в языкоглоточный нерв (IX пара) и далее попадают в ушной ганглий. Его постганглионарные волокна заканчиваются в околоушной слюнной железе.
В состав языкоглоточного нерва входит афферентная синусная ветвь, связанная с большим числом баро- и хеморецепторов каротидного клубочка, располагающегося между внутренней и наружной сонными артериями у места деления общей сонной артерии. От этих рецепторов поступает информация о величине артериального давления крови, pH крови, напряжения в крови кислорода (0 2) и диоксида углерода (С0 2). Афферентные импульсы участвуют в рефлекторной регуляции функций сердечно-сосудистой системы, а также дыхания.
Из ядер слезоотделительного пути продолговатого мозга преган- глионарные волокна в составе лицевого нерва (VII пара) вступают в крыловидный ганглий, постганглионарные волокна которого иннервируют слезные и слюнные железы, железы слизистой оболочки носовой полости и нёба.
В продолговатом мозге находятся ядра, в которых расположены тела нейронов, преганглионарные волокна которых участвуют в образовании блуждающего нерва (X пара). Блуждающий нерв смешанный: он состоит из афферентных и эфферентных парасимпатических, эфферентных симпатических, чувствительных и двигательных соматических волокон. Однако преобладают афферентные чувствительные волокна, которые передают информацию от рецепторов органов грудной полости к органам брюшной полости. Рецепторы реагируют на механические, термические, болевые воздействия, воспринимают изменения pH и электролитного состава внутренней среды организма.
Важную физиологическую роль выполняет ветвь блуждающего нерва - депрессорный нерв, по которому проходит информация, сигнализирующая о функциональном состоянии сердца и величине кровяного давления в дуге аорты. Нейроны ядер афферентных путей блуждающего нерва лежат в яремном узле, а их аксоны проникают в продолговатый мозг на уровне олив. Ганглии находятся в иннервируемом органе или около него.
Передача возбуждения с аксонов первых нейронов (преганглионарные волокна) на нейроны ганглиев и с аксонов нейронов парасимпатических ганглиев (постганглионарные волокна) на структуры органа осуществляется через синапсы с помощью медиатора ацетил- холина.
Преганглионарное волокно более длинное и идет от центральной нервной системы до органа, постганглионарное волокно более короткое.
Значение парасимпатической иннервации. Основная роль парасимпатической нервной системы состоит в регуляции различных функций, обеспечивающих гомеостаз - относительное динамическое постоянство внутренней среды организма и устойчивость основных физиологических функций. Парасимпатическая иннервация обеспечивает восстановление и поддержание этого постоянства, дестабилизированного активацией симпатической нервной системы. Парасимпатические нервные волокна совместно с симпатическими волокнами обеспечивают оптимальное функционирование органов, которые они иннервируют. При активации парасимпатичекой системы проявляются реакции, противоположные действию симпатической нервной системы. Например, это приводит к снижению частоты и силы сокращений сердца, сужению бронхов, активации слюноотделения и т.д.
Оглавление темы "Вегетативная (автономная) нервная система.":1. Вегетативная (автономная) нервная система. Функции вегетативной нервной системы.
2. Вегетативные нервы. Точки выхода вегетативных нервов.
3. Рефлекторная дуга вегетативной нервной системы.
4. Развитие вегетативной нервной системы.
5. Cимпатическая нервная система. Центральный и переферический отдел симпатической нервной системы.
6. Симпатический ствол. Шейный и грудной отделы симпатического ствола.
7. Поясничный и крестцовый (тазовый) отделы симпатического ствола.
9. Периферический отдел парасимпатической нервной системы.
10. Иннервация глаза. Иннервация глазного яблока.
11. Иннервация желез. Иннервация слезной и слюных желез.
12. Иннервация сердца. Иннервация сердечной мышцы. Иннервация миокарда.
13. Иннервация легких. Иннервация бронхов.
14. Иннервация желудочно-кишечного тракта (кишечника до сигмовидной кишки). Иннервация поджелудочной железы. Иннервация печени.
15. Иннервация сигмовидной кишки. Иннервация прямой кишки. Иннервация мочевого пузыря.
16. Иннервация кровеносных сосудов. Иннервация сосудов.
17. Единство вегетативной и центральной нервной системы. Зоны Захарьина - Геда.
Парасимпатическая часть исторически развивается как надсегментарный отдел, и поэтому центры ее располагаются не только в спинном мозге, но и в головном.
Центры парасимпатической части
Центральная часть парасимпатического отдела состоит из головного, или краниального, отдела и спинномозгового, или сакрального, отдела.
Некоторые авторы считают, что парасимпатические центры располагаются в спинном мозге не только в области крестцовых сегментов, но и в других отделах его, в частности в пояснично-грудном отделе между передним и задним рогом, в так называемой интермедиарной зоне. Центры дают начало эфферентным волокнам передних корешков, вызывающих расширение сосудов, задержку потоотделения и торможение сокращения непроизвольных мышц волос в области -туловища и конечностей.
Краниальный отдел в свою очередь состоит из центров, заложенных в среднем мозге (мезэнцефалическая часть), и в ромбовидном мозге - в мосту и продолговатом мозге (бульбарная часть).
1. Мезэнцефалическая часть представлена nucleus accessorius n. oculomotorii и срединным непарным ядром, за счет которых иннервируется мускулатура глаза - m. sphincter pupillae и m. ciliaris.
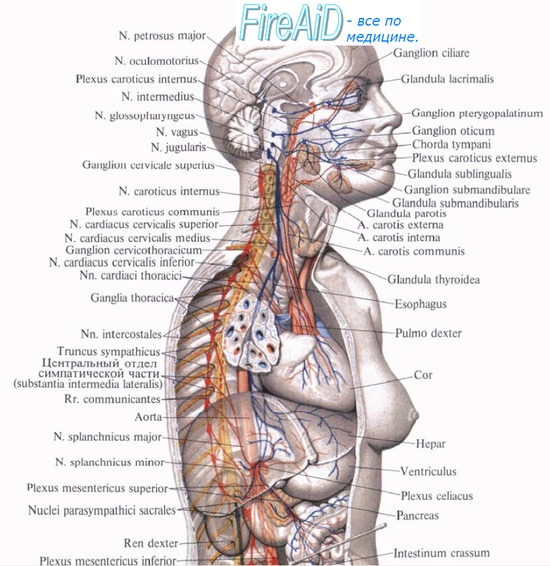
2. Бульварная часть представлена nucleus saliva tonus superior n. facialis (точнее, n. intermedius ), nucleus salivatorius inferior n. glossopharyngei и nucleus dorsalis n. vagi (см. соответствующие нервы).
ВНС делится на два отдела – симпатический и парасимпатический. По строению они различаются расположением своих центральных и эффекторных нейронов, своими рефлекторными дугами. Они различаются так же и по своему влиянию на функции иннервируемых структур.
В чем состоят различия этих отделов?
Центральные нейроны симпатической нервной системы расположены, как правило, в сером веществе боковых рогах спинного мозга от 8 шейного до 2-3 поясничных сегментов. Таким образом, симпатические нервы всегда отходят только от спинного мозга в составе спинномозговых нервов по передним (вентральным) корешкам.
Центральные нейроны парасимпатической же нервной системы находятся в крестцовых сегментах спинного мозга (2-4 сегменты), но большая часть центральных нейронов находятся в стволе мозга. Большая же часть нервов парасимпатической системы отходят от головного мозга в составе смешанных черепно - мозговых нервов. А именно: из среднего мозга в составе III пары (глазодвигательный нерв) - иннервируя мышцы ресничного тела и кольцевые мышцы зрачка глаза, из Варолиевого моста выходит лицевой нерв - VII пара (секреторный нерв) иннервирует железы слизистой оболочки носа, слёзные железы, подчелюстную и подъязычную железы. Из продолговатого мозга отходит IX пара - секреторный, языкоглоточный нерв, иннервирует околоушные слюнные железы и железы слизистой щек и губ, X пара (блуждающий нерв) - самая значительная часть парасимпатического отдела ВНС, проходя в грудную и брюшную полости, иннервирует весь комплекс внутренних органов. Нервы, отходящие от крестцовых сегментов (2-4 сегменты), иннервируют органы малого таза и входят в состав подчревного сплетения.
Эффекторные нейроны симпатической нервной системы вынесены на периферию и находятся или в паравертебральных ганглиях (в симпатической нервной цепочке), или превертебрально. Постганглионарные волокна образуют различные сплетения. Среди них наиболее важное значение имеет чревное (солнечное) сплетение, но в его состав входят не только симпатические, но и парасимпатичесике волокна. Оно обеспечивает иннервацию всех органов расположенных в брюшной полости. Вот почему так опасны удары и травмы верхней части брюшной полости (примерно под диафрагму). Они способны вызвать шоковое состояние. Эффекторные нейроны парасимпатической нервной системы всегда находятся в стенках внутренних органов (интрамурально). Таким образом, у парасимпатических нервов большая часть волокон покрыты миелиновой оболочкой, и импульсы достигают эффекторных органов быстрее, чем у симпатической. Это обеспечивает парасимпатические нервные влияния, обеспечивающие сбережение ресурсов органа и организма в целом. Внутренние органы, расположенные в грудной и брюшной полости иннервируются главным образом блуждающим нервом (n.vagus), поэтому эти влияния часто называют вагусными (вагальными).
Имеются существенные различия и в их функциональных характеристиках.
Симпатический отдел, как правило, мобилизирует ресурсы организма для осуществления энергичной деятельности (усиливается работа сердца, сужается просвет кровеносных сосудов и повышается артериальное давление, учащается дыхание, расширяются зрачки и т.п.), но происходит торможение работы пищеварительной системы, за исключением работы слюнных желез. У животных это происходит всегда (слюна нужна им для зализывание возможных ран), но и у некоторых людей при возбуждении слюноотделение усиливается.
Парасимпатическая, напротив, стимулирует работу пищеварительной системы. Неслучайно после сытного обеда отмечается вялость, нам так хочется поспать. При возбуждении парасимпатической нервная система обеспечивает восстановление равновесия внутренней среды организма. Она обеспечивает работу внутренних органов в состоянии покоя.
В функциональном смысле симпатическая и парасимпатическая системы являются антагонистами, дополняя друг друга в процессе поддержания гомеостазиса, поэтому многие органы получают двойную иннервацию - и со стороны симпатического, и со стороны парасимпатического отделов. Но, как правило, у разных людей преобладает или тот или другой отдел ВНС. Неслучайно известный отечественный физиолог Л.А. Орбели попытался классифицировать людей по этому признаку. Он выделил три типа людей: симпатикотоники (с преобладанием тонуса симпатической нервной системы) - их отличает сухость кожи, повышенная возбудимость; второй тип - ваготоники с преобладанием парасимпатических влияний - для них характерна жирная кожа, замедленные реакции. Третий тип -промежуточный . Л.А. Орбели считал знание этих типов важным для врачей, особенно при назначении доз лекарственных препаратов, т. к. одни и те же лекарственные препараты в одинаковой дозе по-разному влияют на пациентов с разным типом ВНС. Даже из повседневной практики каждый из нас может заметить, что чай и кофе вызывают различную реакцию у людей с разным типом функциональной активности ВНС. Из экспериментов на животных известно, что у животных с разным типом ВНС введение брома и кофеина так же оказывает различные реакции. Но на протяжении жизни человека его тип ВНС может изменяться в зависимости от возраста, периода полового созревания, беременности и других влияний. Несмотря на перечисленные различия, обе эти системы, однако, составляют единое функциональное целое, т. к. интеграция их функций осуществляется на уровне ЦНС. Вы уже знаете, что в сером веществе спинного мозга центры вегетативных и соматических рефлексов успешно соседствуют, также как они располагаются близко друг с другом в стволе мозга, и в высших подкорковых центрах. Так же как, в конечном счете, в единстве функционирует вся нервная система.
Подкорковые высшие центры ВНС находятся в гипоталамусе, который связан обширными нервными связями с другими отделами ЦНС. Гипоталамус является в то же время частью лимбической системы мозга. Функции вегетативной нервной системы, как известно, не контролируются сознанием человека. Но именно через гипоталамус и (связанный с ним гипофиз) высшие отделы ЦНС способны влиять на функциональную активность вегетативной нервной системы и через неё на функции внутренних органов. Функции дыхательной, сердечно-сосудистой, пищеварительной и других систем органов непосредственно регулируются вегетативными центрами, расположенными в среднем, продолговатом отделах головного и отделах спинного мозга, которые подчинены в своих функциях центрам гипоталамуса. В то же время туда же продолжаются ядра черной субстанции, черные ядра, располдоженные и в среднем мозге, ретикулярная формация. Действительно, реализация влияния психических реакций человека на соматические – повышение артериального давления при гневе, повышенное потоотделение при страхе, пересыхание во рту при волнении и многие другие проявления психических состояний, – происходит при участии гипоталамуса и ВНС под влиянием коры больших полушарий.
Гипоталамус является частью промежуточного мозга. В нем можно выделить передний отдел (передний гипоталамус) и задний отдел (задний гипоталамус). В гипоталамусе расположены многочисленные скопления серого вещества - ядра. Их более 32 пар. По своему расположению они делятся на области - преоптическую, переднюю, среднюю и заднюю. В каждой из этих областей лежат группы ядер, отвечающих за вегетативную регуляцию функций, а также ядра, выделяющие нейрогормоны. Эти ядра различают также по их функциям. Так, в передней области находятся ядра, выполняющие функции регуляции теплоотдачи за счёт расширения кровеносных сосудов и увеличения отделения пота. А ядра, регулирующие теплопродукцию (за счёт повышения катаболических реакций и непроизвольных мышечных сокращений), располагаются в задней области гипоталамуса. В гипоталамусе расположены центры регуляции всех видов обмена веществ - белкового, жирового, углеводного, центры голода и насыщения. Среди групп ядер гипоталамуса находятся центры регуляции водно-солевого обмена, связанные с центром жажды, формирующего мотивацию поиска и потребления воды.
В передней области гипоталамуса лежат ядра, участвующие в процессах регуляции чередования сна и бодрствования (циркадных ритмов), а так же в регуляции полового поведения.
Проекции вегетативных центров представлены и в коре больших полушарий - в основном в лимбической и ростральной части коры. Парасимпатические и симпатические проекции одних и тех же органов проецируются в одни и те же или близко расположенные участки коры, это понятно, т. к. они совместно обеспечивают функции этих органов. Установлено, что парасмпатические проекции в коре представлены гораздо шире, чем симпатические, однако, функционально симпатические влияния более продолжительны, чем парасимпатические. Это связано с различиями медиаторов , которые выделяются окончаниями симатических (адреналин и норадреналин) и парасимпатических (ацетилхолин) волокон. Ацетилхолин - медиатор парасимпатической системы - быстро инактивируется ферментом ацетилхолинэстеразой (холинэстеразой) и её влияния быстро сходят на нет, в то время как адреналин и норадреналин инактивируются значительно медленнее (ферментом моноаминоксидазой), их влияние усиливается норадреналином и адреналином, выделяемыми надпочечниками. Таким образом, симпатические влияния длятся дольше и оказываются более выраженными, чем парасимпатические. Однако, во время сна парасимпатические влияния на все наши функции превалируют, что способствует восстановлению ресурсов организма.
Но, несмотря на различия в строении и функциях различных отделов ВНС, различия соматической и вегетативной систем, - в конечном итоге, вся нервнаяя система работает как единое целое и интеграция происходит на всех уровнях как спинного, так и головного мозга. И высшим уровнем интеграции, безусловно, является кора больших полушарий головного мозга, объединяющая как нашу двигательную активность, работу наших внутренних органов так и, в конечном итоге, всю психическую деятельность человека.
18. Физиология надпочечников, роль их гормонов в регуляции функций организма, взаимоотношении с другими регуляторными механизмами .